Macroscopic:
To understand what the microscopic (molecular) reasons are as to why different species age at different rates one first has to normalize their (maximum) lifespans in respect to the macroscopic parameters temperature and mass specific energy turnaround.
As a first approximation one gets the result that the maximum lifespan is inversely proportional to the mass specific energy consumption (neglecting temperature for the time being). Or put it differently, the product of the maximum lifespan and the mass specific energy expenditure is constant. This is the so called Rate Of Living Theory of Ageing. It says that 1 gram of biological matter can burn roughly 500 kcal before it is exhausted. For the whole organism one can therefore calculate a so called life potential which is the number of calories it can burn until it inevitably dies.
Ageing therefore is a general property of biological matter and is not due to the specific construction of a certain species (e.g. genes), at least as a crude approximation.
A better understanding of this theory has been given by West et al.,
see e.g. Biological Scaling and Physiological Time and the lecture 1 2 3 4 5 6 7 8
Their claim is that
![]()
The quarter power dependence of the maximum lifespan is due to a geometric effect. Qualitatively it can be understood quite easily, though it is a bit bit difficult to derive mathematically.
It has to do with the fact, that energy is supplied via so called branching networks, the blood circulary system being one example. Naively one would expect that if one doubles the mass of an organism the energy supply capacity (e.g. the mass of the capilaries) would also double for the individual cell to receive the same amount of nutrients and energy. Surprisingly this is not the case, as these branching networks are fractal in nature and their mass scales as m^0.75. This means that the bigger an animal is, the less energy an individual cell can maximally receive and the higher the upper limit for the maximum lifespan is. (Whales being a suggestive example). The maximally possible energy expenditure therefore scales as m^0.75.

To test this hyothesis I have ploted data, taken from AnAge database which is a great source of ageing related data.
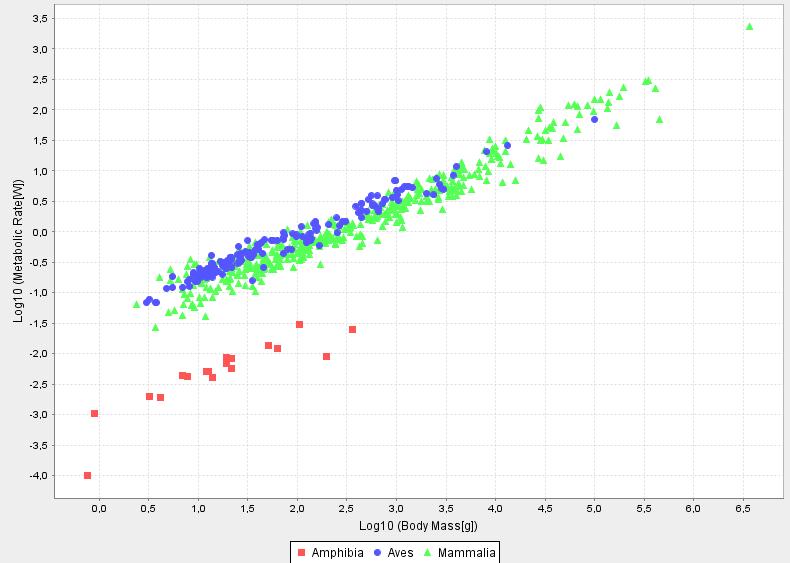
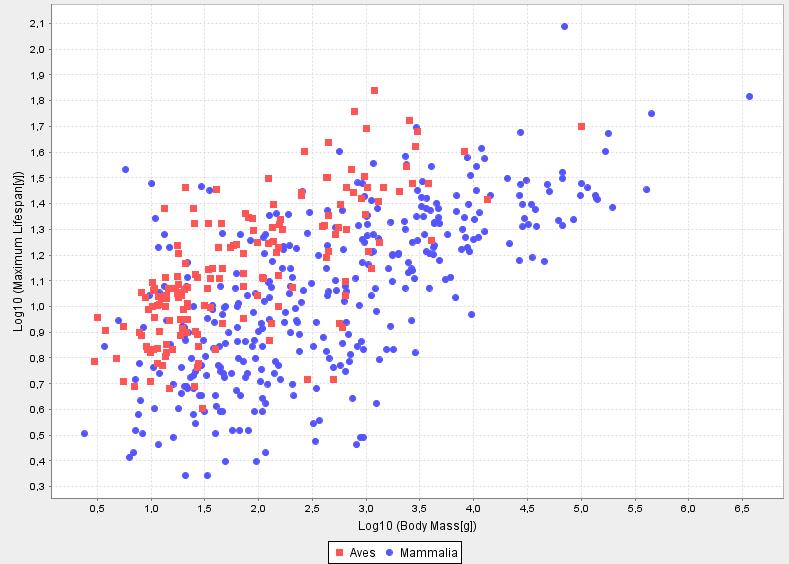
If the rate of living theory where exactly fulfilled, L0 would be the same for all species. This is not the case, still the rate of living theory works remarkably well and in my opinion can well serve as a paradigm for further investigations (i.e. trying to understand why it is approximately fulfilled so well and why deviations occur for certain species). Outstanding exceptions to the theory are for example birds and bats who live exceptionally long.
To further investigate the microscopic differences of ageing between different species one can define
L0 = M*LR.L.
with L0 see above, LR.L. the rate of living constant and M a factor that I call "maintenance factor" which describes the deviations from the rate of living theory. Birds and bats for example would have a maintenance factor larger than 1 as they live longer than average.
The following plot shows maintenance factors for different species (data from AnAge) which shows that there clearly are molecular differences between different phyla that contribute to a different rate of ageing:
... TODO insert plot ...
Interesting in the context of the rate of living theory is a study done with birds: Tropical Birds Have Slow Pace Of Life Compared To Northern Species, Study Finds
The answer to the question what the reason for these microscopic differences could be will be persued under "microscopic".
Reliability Theory:
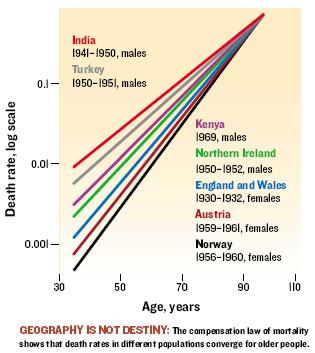
Mortality curves (i.e. plots of mortality vs. time) show a universal behaviour which is the so called Gompertz law and which is a special form of a logistic probability distribution. These distributions are based on reliability theory, i.e. the body has a redundancy in its subsystems that fail one after another as time passes leading to a system's failure once a critical treshold is reached. (Interestingly failure rates of light bulbs also show a logistic behaviour).
The convergence of the death rates reflects the fact, that there is a (universal) maximum lifespan for a given species. The different curves for different populations correspond to different mean life spans ("rectanglification of the mortality curve").
- A slow down in ageing should manifest itself in a right-shift of the mortality curve, i.e. an increased maximum lifespan. Interventions that yield this are the real anti-ageing interventions.
- An increase in the mean lifespan, leaving the maximum lifespan unchanged, just means a decrease in the number of premature deaths, i.e. a less "suboptimal" ageing.
Microscopic:
Ageing is a general phenomenon and not restricted to biological systems and can be described as loss of information of a system with time.
The pacemakers of ageing in biological systems therefore must be structures with lifetimes of the order of the lifetime of the species that serve as the backup structures for the information. Candidates are DNA (+ methylation patterns) and mtDNA. RNA cannot serve as backup structures as due to the central dogma of biology information flows from DNA to RNA only. Furthermore unwanted stable structures, that gradually build up during lifetime and "seed misinformation" and hinder processes also have to be considered (AGEs and other kind of junk). These stable structures therefore can be seen as "ageing clocks", that is if one looks at them one should be able to determine its biological age. They therefore could serve as biomarkers for ageing.
One of the questions in search for a microscopic theory of ageing is: Is there one key effect or are there a small number of key effects that are the pacemakers of ageing, or is ageing a superposition of many effects. If the latter is the case, I think it will be very difficult to fundamentally counteract ageing. I suggest a theory for this below ("Theory of quasi synchronous ageing").
The most prominent theory at the moment in the scientific community alluding to the former situation, i.e. there being a small number of "bottlenecks" or one single "bottleneck", preventing a considerably longer lifespan, seem to be the free radical theory of ageing. Closely related to it is the mitochondrial theory of ageing.
Free Radical Theory of Ageing:
